Instructions And Tips For Growing FPsc
Instructions_and_Tips_for_Growing_FPsc
Instructions_and_Tips_for_Growing_FPsc
Instructions_and_Tips_for_Growing_FPsc
User Manual: Pdf
Open the PDF directly: View PDF ![]() .
.
Page Count: 18

Instructions and Tips for Growing Rapid-cycling Brassica rapa variety “FPsc”
Brassica rapa FastPlants sc (FPsc) is a plant variety specifically bred for educational use in classrooms
and laboratories. It is outwardly similar in gross phenotype and growth habit to the famous Wisconsin
Fast Plants (WFP) variety of rapid-cycling B. rapa, but differs in ways that enable teachers and students
to take an integrated approach to Mendelian and molecular genetics. Thus, like WFP, FPsc is rapid-
cycling, flowering ~18 days after planting. The plants are relatively compact and need very little inputs or
maintenance to complete the life cycle (seed to seed) in ~56-60 days. The essential difference between
FPsc and WFP is that FPsc is self-compatible (hence the “sc” in its name) and tolerates inbreeding (via
self-pollination, or ‘selfing’) with no consequent inbreeding depression. FPsc has been extensively
inbred: seeds prepared for distribution were derived from a plant lineage that had progressed through 7
generations of single-seed descent by selfing. Since in each such generation the residual heterozygousity
is reduced by ½, we estimate that FPsc is homozygous at >99% of all loci in the genome. Absent allelic
variation, pure (i.e., non-hybrid) FPsc stocks can readily be propagated over successive generations and
still maintain the original genetic constitution—i.e., FPsc is true-breeding.
Growing FPsc is little different than growing WFP—add seeds to wet soil, put them under the lights and
let them grow. Many useful tips for growing B. rapa and classroom activities can be found at the WFP
home page (http://www.fastplants.org/) and nearly all are applicable to propagation of our FPsc variety.
However, there are several tips and observations we’ve made that may be useful to growers to enable
optimal growth, seed yield, and classroom performance. In the following we provide detailed
instructions for growing FPsc. The directions are in standard font, tips and background information are
provided in italics.
We thank you for your interest in using these exciting new resources. Please do not hesitate to email
(swoody@wisc.edu or amasino@biochem.wisc.edu ) if you have any questions or would like additional
information.
Scott Woody and Rick Amasino
University of Wisconsin-Madison
September, 2011

Propagation of FPsc
1. Place soil in the growing vessel and wet thoroughly with water.
Soil: FPsc grows well in a variety of commonly available soil mixes. We routinely use MetroMix
360 but have had good success using alternative soils, including ReadiEarth, Fafard #3, and even the
Miracle Grow potting soil (available at most garden stores). These soils require only a light supplement
with a balanced fertilizer (e.g., Peters 10-10-10, Osmocote pellets, or equivalent). It is important that
micronutrients/trace elements be provided in the soil or through fertilizer supplement. It can be difficult
to know in advance whether the soil you select contains micronutrients. For example, MetroMix 360 and
MetroMix 300 products have essentially the same ingredients listed by the manufacturer. However, our
plants grow as if starved on MetroMix 300 but need no supplemental nutrients to complete the life cycle
if grown on 360. Whatever soil you use, it helps to make sure that it is uniform with respect to texture.
Obvious clumps should be broken-up or discarded (figure 1). Fill the vessel lightly to avoid compacting
the soil and create a level surface when complete.
Figure 1: Try to ensure that your soil is of uniform consistency. Break up clumps and
discard large chunks of bark and sticks. Our goal in genetic analysis is to compare
differences in phenotype that are due only to the activity or effect of alternative
alleles; we want all other environmental conditions to be as uniform as possible.
Growing vessel: This is a clumsy way of saying “pot”, but allows that our plants will grow well in
a variety of containers. We routinely grow plants in cells of plastic inserts like those used by garden
stores to sell marigolds, tomatoes, etc. (figure 2). As a general rule, the larger the volume of soil
provided, the larger and more vigorous will be the plants, although there are limits at both extremes—at
some point adding more soil volume has no further effect on plant size or vigor and very small volumes (<
50 cm3) should be avoided.

Figure2: Recommended growing configuration. Plastic inserts (72-cell, left, or 48-
cell, right) are placed in a shallow reservoir to which water will be added to irrigate
the soil. Both configurations provide easy division (using pre-cut perforations) of
plant populations into convenient units for distribution to teams of students. Fill
with uniform and fluffy soil and level the surface prior to watering and planting.
Wet thoroughly: To promote rapid and uniform germination, it is important to create a
microenvironment around the seed that is at or near saturation with respect to humidity/moisture. In
subsequent steps we will describe additional steps we recommend to achieve this condition; for present
purposes we note only that the soil should be thoroughly wet prior to planting but there should not be
standing water—be sure that the vessel you use has drain holes in the bottom to release excess water
from the soil. A good way to wet the soil is to use a gardening pail with a diffusing spout. You want to
create a gentle “rain shower” that wets the soil without leading to compaction or disruption of a
uniform, level surface.
Water: We use regular tap water from the university supply lines but this may not be suitable in
all locations or at all times of the year. If you have concerns, use distilled/deionized water if possible.
2. Dispense seeds into the palm of your hand or onto a seed tray and place seeds individually
on the surface of the wet soil.
Seed tray: We typically use 4” x 6” note cards with all edges folded up ~3/4” to prevent the seeds
from rolling away (figure 3).
Figure3: Seed tray made from a note cards. Seeds may be planted by hand, with the
use of fine tip forceps, or by using the jagged and wettened end of a broken large
toothpick or bamboo skewer.
Place seeds: Both terms merit comment. To “place” the seeds means to deposit them gently on
the soil surface. There is no need (and it may be disadvantageous) to press the seeds down into the soil.
If your fingers are small and nimble, use them; alternatively, you can use forceps but be careful—
depending on the style and width of the forcep tips seeds can sometimes “pop” loose and land (a) where
you can’t find them and/or (b) precisely where you don’t want them. A third option is to use the jagged
tip a broken toothpick or bamboo skewer that has been wetted with water or saliva. Capillary action
holds the seed to the wooden edge during transit but releases the seed easily when it is placed onto wet
soil.
Seeds: The quality of seeds that are planted is among the most important factors that determine
the vigor of the resulting plants. Embryogenesis/seed development in FPsc and many other varieties of B.
rapa is “messy”—some proportion of fertilized ovules abort development altogether at some point
during embryogenesis; in others, development proceeds slowly and with apparent difficulty, resulting in
undersized and sometimes misshapen embryos/seeds at maturity. The cause(s?) of spontaneous
abortion or stunted embryo development are unknown; in any case, the proportion of “bad seeds” is
higher in plants grown under sub-optimal conditions. “Good seeds” are full and round (potato-shaped,
actually; see fig. 4), whereas the seed coats of “bad seeds” are typically shrunken and/or wrinkled, an
outward manifestation of the embryo within. Many of the latter can germinate but the resulting plants
are frequently and likewise misshapen, and overall vigor is typically reduced as compared to plants
grown from “good” seeds. We have a mantra in the lab that goes something like “Crappy plants make
crappy seeds and crappy seeds make crappy plants”. We try to package only the highest quality of seeds
for distribution, but even after our efforts there may be variation in seed quality. Furthermore, if your
experiments span multiple generations, you need to be aware of bad seeds. In any case, it is important to
select the best seeds available when planting and to grow plants under the best possible conditions in
order to maximize the proportion of high quality seeds.
[It’s worth noting that most types of B. rapa “parts” you may see in the veggie section of the
supermarket are the product of selective breeding programs imposed by humans, likely conducted over
many thousands of years and seed production has rarely been a driving force since the portions that are
intended to be eaten are produced during the vegetative phase of plant development. Think of the
turnips (B. rapa var. rapifera); the fleshy leaves of Bok choi (B. rapa var. pekinensis) and Pak Choi plants,
and the spicy edge provided by the thin and serrate leaves of Chinese mustard greens (B. rapa var.
Mizuna). It’s all about the leaf, and as long as seed –production is sufficient to provide for next season’s
crop, maximizing seed yield and seed quality has not been as direct a target for selection by those
breeders. An exception to this general rule are B. rapa oil seed crops grown in northern US and
international latitudes (rapeseed) and in Asiatic countries such as India, China and Nepal, where the
seeds are an important source of spices and condiments.

Figure 4: Plant the best seeds that you have available—size matters, and bigger is
better. Try to avoid shrunken or otherwise misshapen seeds, as well as any whose
embryos had bypassed dormancy and started to germinate (vivipary).
3. Once all seeds have been placed/planted, sprinkle coarse grade vermiculite or additional potting soil
over the seeds and soil surface to a thickness just sufficient to cover both seeds and soil. Using the
watering pail with a diffusing/sprinkler head, lightly add water to thoroughly wet the vermiculite.
Vermiculite is available at most horticultural supply stores or web sites. We use Sunshine coarse
grade (Sun Gro Horticulture, Bellvue, WA), but largely out of habit. Alternative brands should be equally
suitable. Vermiculite is a mined and processed mineral similar in terms of multilayered structure to mica.
Upon heating, microdroplets of water trapped between the layers turn to steam, causing the mineral to
expand as the layers separate slightly. When used for growing our plants, vermiculite serves two
purposes: first, upon wetting, water is drawn between layers of the mineral and held in place by capillary
action. With the wet soil beneath and the wet vermiculite around and above, the seeds are placed in a
very moist—but not suffocating—environment that promotes rapid germination. Second, by shielding
the organic material in the soil from light, the vermiculite layer limits or prevents growth of algae during
the plant life cycle.
Figure5: Magnified (6x) view of highly-layered vermiculite (left) and a covered flat
after planting seeds (right). Gently water the flat from above to moisten the
vermiculite. If vermiculite is unavailable, a light layer of potting soil may be used to
cover seeds.
4. Place the flat, pot, or other container under lights in an area where temperatures can
be maintained 18-24 degr. C. Prop up the pots (or lower the lights) to leave ~10 cm (4”)
between lights and the soil surface or (later) the apex of the plants.

Lights and distance: Here at the UW-Madison, we have the luxury of growing plants in controlled
environmental chambers. Specifically, our plants are grown in 11” x 22” flats placed beneath 4’, 4-bulb
32 watt T8 cool white florescent fixtures. The room temperature is held at or near 22 degr. C throughout
the plant life cycle and lighting is continuous—i.e., no light/dark cycles are imposed. Flats are supported
by an 11” x 22” sheet of ¼” plexiglass which is, in turn, elevated by placement of empty pots or flats to
maintain a distance between plants and bulbs of ~10 cm; supports beneath the plexiglass are altered or
removed as plants grow taller in order to maintain that approximate distance (figure 6).
Figure6: Freshly planted and watered flats under lights. Lower supports are
removed as plants emerge and grow; we aim to keep the apices ~4 inches from the
light.
We have examined growth of FPsc under a variety of lighting and temperature conditions. Under
the conditions described above, light incidence ~4” from the bulbs is ~9,000 lux (~900 ft-candles). For the
sake of comparison, light intensity at the benchtop in our lab measures ~900 lux; natural light coming in
the window on an overcast Wisconsin day in late September is ~3500 lux and ~37,000 lux in full 3 p.m.
(late September) sunlight. In general, more light is better than less. For example, we have 55 watt T5HO
fixtures in one of our growth rooms. Plants grown under those lights receive ~35,000 lux at 4” and show
no ill effects. On the other hand, insufficient light can lead to problems. In particular, seedlings grown
under cool white lights at a distance that yields ~5,500 lux show modest signs of etiolation (extended
hypocotyls) and slight chlorosis (they are a bit paler green than seedlings grown with 9,000 lux).
Ultimately, plants grown under conditions of insufficient light are spindly and weak.
A simple experiment to test whether your setup provides sufficient light intensity (and an
interesting one for students to do) is to allow seeds to germinate in pots placed at variable distance from
the light source(s) and measure hypocotyl length at ~ 1 week post-germination. You will notice that
hypocotyl lengths will increase in proportion to the distance from the lights (intensity decreases as the
square of distance). The objective is to find the distance/intensity at which no further decrease in
hypocotyls length is observed as compared to plants grown under higher intensity (achieved by moving
plants closer to the light). In our hands, 10,000 lux provides saturating light intensity but note that you
don’t need a meter to determine if your setup is sufficient for good growth.
A couple more things on lighting are worth noting. First, there are many inexpensive lighting
systems on the market—Carolina Biological Supply is a good source, as are the many horticulture sites
online-- or you can rather easily build your own. Second, if you find that your lighting is not sufficiently
intense, you may need to change the bulbs. Output from florescent lamps decreases over time, so new
bulbs may be a quick and cheap fix. Finally, don’t bother with “grow lights”. Whereas they may be useful
for propagation of certain plant varieties, we have found that they provide no discernable benefit for
growth of B. rapa.
Temperature: FPsc grows well in a temperature range of 15 – 25 degr C. At temperatures
towards or below the low end of that range growth is notably slower, and signs of stress and wilting are
evident at or above 25 degr. C.
5. The seeds should germinate within a day or two and you will see the hypocotyl and cotyledons
emerge from the layer of vermiculite. Keep the soil moist but not saturated during plant growth.
Depending on the soil mix that you use, a supplemental feeding at about one or two weeks may be
useful but don’t overdo it with the fertilizer. Plants should make 2-4 true leaves and then transition to
flowering. Typically the first flowers are open at ~18 days after planting.
Keep the soil moist but not saturated: FPsc (and most other non-wetland plants) does not grow
well in water-saturated soil. The roots need to breath and chlorosis or death will result if the plants are,
in essence, drowning. There are two ways we know of to maintain optimal soil moisture levels: wicking
and periodic flooding. Wicking is widely used in classrooms for propagation of the WFP variety of B. rapa
and also works well for growing FPsc. Briefly, a segment of yarn, cotton string, or other absorbamt and
water-conducting material is inserted in the pot before soil is added, threaded out the bottom and into a
reservoir of water. Water wicks up the fibers and into the soil as transpiration and evaporation remove it
during plant growth. Use of a wicking strategy imposes front-end costs since it takes time to wick each
pot or cell but those costs may be recovered later in terms of vigorous growth and less need to constantly
monitor soil moisture levels. The latter is especially useful in classroom settings since teachers (and
students) can leave for the weekend and be confident that the plants will be well when classes resume
(provided, of course, that the reservoir was topped-off with water before leaving). Periodic flooding is
another useful approach to maintaining optimal moisture levels. The pots with soil and plants are
immersed in a larger vessel filled with water to a level sufficient to saturate the soil. Briefly flooded, the
flats/pots are removed and excess water is allowed to drain before moving plants back to lighted
shelves. Again, here at the UW, we have the luxury of access to flood tables in which electronically-
controlled water pumps actuate to flood and then drain the flats thrice weekly, but we use those
facilities only when we are bulking seed stocks or otherwise growing exceptionally large numbers of
plants. It would be rather painful if we used the approach for all of our experiments since it can be messy
and it takes time. Instead, we settle for a middle ground approach, adding water to the white flat
bottom (Fig. 1) in small doses several times per week, providing somewhat bigger “drinks” if we know
that plants will be unattended for 2-3 days.

Please also be aware that water usage by your plants will vary as a function of developmental
stage, reaching a maximum at or about the time of flowering and seed set when there is an abundance
of photosynthetically active leaves that both consume water and serve as platforms for water loss
through transpiration and evaporation.
Supplemental feeding: As noted in item 1, the extent to which supplemental fertilizer/nutrients
are needed depends heavily on which soil mix is used. Also as noted, we have found that MetroMix 360
has sufficient resources to support good growth without any supplemental feeding; nonetheless, we
typically feed them lightly with a balanced fertilizer containing trace elements/micronutrients just before
the onset of flowering. We use Peters All Purpose plant food (20-20-20 with micronutrients) at ½
recommended strength and added instead of plain water when the flat bottoms/reservoirs are dry (or
nearly so). As with many things, less is more. Paul Williams (father of the WFP program) once told me
that his goal when growing B. rapa is to keep them adequately nourished but “just a little hungry”. Too
much fertilizer can be detrimental in (at least) two ways. First, if added at high rates at planting,
especially if added to a soil mix already endowed with fertilizer, salts in the soil may come out of solution
and crystallize on plant surfaces. In extreme cases this leads to what we call ‘tip burn’, in which seedlings
emerge, spread their cotyledons, and then growth stalls due to damage at the shoot apical meristem.
Often plants suffering tip burn never resume growth and ultimately die; alternatively, cells of the apical
meristem may reorganize to permit further growth but such meristems are apparently dysfunctional—
leaves that emerge are often misshapen, we see fasciation of the stem, etc (figure 7). Those plants are
clearly not well, and remember the maxim: crappy plants make crappy seeds, etc. Thus, if you suspect
that your soil may be deficient in minerals and nutrients, wait about a week until the seedlings have
emerged and the meristems are elevated from the soil surface to minimize contact with any crystallized
salts. The second detrimental effect of excess fertilizer is the tendency of overfed plants to produce
secondary branches and to continue flowering well past some optimal point (that optimum will be
discussed in item 6, below). The net result is a population of plants that are intertwined due to branching
and slow to reach maturity and senescence.
Figure7: Seeds will germinate and grow above the soil after ~3-4 days (left: seedlings
of a family in which the albino mutant allele is segregating). Center: A stunted
seedling with a condition known as “tip burn”, caused either by crappy seeds and/or
excess fertilizer and salts that have leached from the soil and damaged tissues that
comprise the shoot apical meristem. Immature floral inflorescences should be
evident 2 weeks after planting (right) and ready for pollination in ~18-20 days.
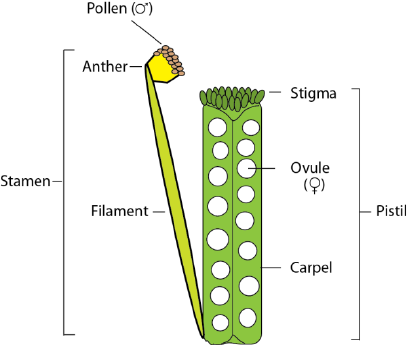
6. Flowering and seed production—general recommendations and comments; instructions for self-
pollination and crosses.
General recommendations and comments: FPsc plants commence flowering at about 18 days
after planting/germination. Flowers are comprised of 4 concentric whorls of discrete organs: 4 sepals
enclosing 4 symmetrically arranged bright yellow petals, 6 stamens (4 long and 2 reduced) and a central
pistil (itself comprised of 2 carpels; figure 8). Healthy plants produce ~3 new flowers/day and flowering
proceeds vigorously for ~5-7 days under standard growing conditions. Pollen grains produced and shed
from the distal ends of stamens (the anthers) will germinate if deposited on the stigmatic cells of the
pistil of the same or other FPsc plant. 15-25 haploid ovules reside within the interior of the pistil;
germinated pollen forms fertilization tubes that burrow through pistil tissues, find and invade the ovules,
and deposit gametes to effect fertilization. Embryogenesis/seed development ensues and is complete in
3-4 weeks. During the course of seed formation, the carpels elongate and enlarge radially to
accommodate the growing embryos. Small, lumpy protrusions will become evident indicating the
location of incipient seeds. Approximately 3 weeks after fertilization, leaf tissues of the plant will show
signs of senescence, losing their previous green color and turgor. Initially the carpels remain green and
are likely the sole site of photosynthesis during late stages of the plant life cycle. Ultimately, however,
the senescence program spreads to the carpels. Initial signs are browning of the “beak”—the pointy tip
of nearly mature seed pods. That browning soon spreads through the carpels as well. At approximately 5
weeks from the onset of flowering, FPsc plants are fully brown and, well… dead. Seeds may be harvested
by cracking dried pods open and capturing in a suitable container.
Figure 8: Schematic representation of floral reproductive organs (sepals and petals
in the outer whorls not shown).
Self-pollination: The FPsc variety is readily selfing. Left unattended, FPsc plants attain ~50% of
the maximal seed yield that can be realized through supplemental hand-pollination. This capacity, an
intended outcome of our breeding program, was enabled by selection early in the program for lines that
had lost (or otherwise did not inherit) the genetic self-incompatibility mechanisms carried by some of the
founding lines. However, there is more to self-fertility than mere absence of self-incompatibility;
developmental and anatomical aspects of flowers are important, too. In most naturally-occurring plant
lines that reproduce exclusively via self-pollination, evolution has produced a well-choreographed
developmental dance between male and female reproductive parts (stamens/anthers and pistils, resp.).
In the model plant species Arabidopsis, for example, the stamens grow adjacent to the pistil but are
slightly longer early in flower development and thus position the anthers (the site of pollen production)
just above the stigma atop the pistil. Furthermore, the pollen-shedding (or dehiscent) surface of the
anthers faces inwards. As the pistil ultimately outgrows the anthers, the stigma pushes through erupting
masses of pollen grains, easily leading to fertilization of all ovules within the pistil. The FPsc variety lacks
such elegant choreography—sometimes the anthers face inwards, sometimes out. The close proximity of
anthers and pistil ensure that some pollination will take place spontaneously, but to ensure maximal
seed production it is helpful to hand-pollinate. Your students will also benefit by close examination of
floral anatomy and the mechanics of pollination.
Hand pollination is pretty simple, but some additional background on plant reproductive biology
might be useful to consider before giving you the ‘how-to’; it may be useful to have a flowering plant to
look at or you can refer to the figures. First, notice that flowering proceeds with sequential maturation
and opening of successive flower buds. In our B. rapa line, 2-3 flowers open per day. Now also notice that
the oldest flowers—the ones that opened earliest, are ‘left behind’ on the growing inflorescence stem. So
regular is the pattern of flowering that merely by counting “down” from the most recently opened
flower, one can estimate how old a particular flower is simply by noting its relative position on the stem.
The last thing you need to know is that pollen grains are not indefinitely viable under normal growing
conditions. For instance (and this is admittedly offered with no evidence but is probably true), it’s likely
that exposure to the air and ambient temperature causes dehydration and “death” of pollen grains in a
fairly short time after flowers open. Alternatively, or in addition, pollen grains are actually autonomous
haploid organisms whose developmental program may have evolved to proceed to senescence and death
unless germination is the triggered by contact with receptive cells atop a compatible pistil. In any case,
pollen grains are mostly unviable by about 4 days after anthesis (the ‘ripe’ stage when flowers are freshly
opened). Finally, and somewhat contrary to the preceding, we have found that pollen grains can retain
viability for several (4-6) months if protected from dessication and stored at low temperatures. We
occasionally take advantage of this property in our experiments—for example, to store pollen from a
flowering plant in order to conduct pollination at a later date. We use a bee stick to gather pollen, place
it in a small sealable tube and store it in the refrigerator until needed.
So now, the ‘how-to’: using a pair of fine-tip forceps, grasp around the base of a 2-3 day old
flower. Holding the stem between thumb and forefinger of one hand, squeeze it near the base and pull to
remove it from the plant. With any luck that gentle squeeze should cause the stamens to splay outwards
from the pistil, exposing the anthers (which should have fluffy mounds of pollen on them). Invert the
detached flower and simply touch the anthers to pistils of successively younger open flowers. If you look
closely, you should see clumps of pollen that were deposited on the stigmas. If you can’t, you may have
picked too-old a flower so try again. With a little practice you’ll find that a single flower can ‘service’ 5-6
younger ones (keeping in mind that your pollinating flower is probably picking up about as much pollen
as it deposits along the way, thereby replenishing its supply).

Crosses: In principle, crosses are no different than self-pollination—the goal remains to deposit
pollen from a given source plant “father” onto the pistil of the intended “mother” plant. The devil is in
the details, so let’s cover those first. The biggest challenge in crossing with FPsc is to avoid inadvertent
self-pollination. This is possible through emasculation (removal of immature anthers) from young flowers
but that raises the second biggest challenge: to avoid killing flowers in the course of emasculation.
Theory and mechanics of successful emasculation. The essential task is to remove some or all
of the floral organs that surround the innermost whorl (the carpels) without damaging the pistil itself.
Specifically, you want to remove the pollen-bearing stamens but it’s simpler and perhaps even necessary
in doing so to remove parts or all of the sepals and petals. The general approach is to use fine-tip forceps
to sever outer whorl organs until enough are removed and you are able to excise the immature anthers.
Thus, identify the oldest unopened flower; grasp the inflorescence stem just beneath the cluster of
immature flower buds and twist gently, if necessary, to give yourself good access to the targeted flower.
It’s useful at this point to remove some of the adjacent and older flowers on the same plant. The end
goal of emasculation is to engineer an exposed pistil ready for pollination by your intended father plant
and you don’t want to risk inadvertent self- or cross-pollination by grains drifting in from other sources.
Figure9: Initial steps in emasculation of flowers. Note—sharp and straight forceps make
the job much easier. Take care to use a Kleenex or other absorbent material to clean
forceps tips prior to emasculation in order to avoid introduction of unwanted pollen
grains.
Now here’s where it gets a bit tricky. Push the straight, sharp tip of your forceps against the
flower. You should aim to have the tips just flank the lateral edges of a sepal at a position ~1/3 of the
way from the base of the flower; higher doesn’t hurt but will likely limit access in later steps and require
more ‘bites’. Lower is BAD since all organs are joined at the base and you risk damaging the pistil in the
next step. Flex the tips to gain some degree of purchase on the sepal and perhaps more interior organs—
not too far, don’t pierce the pistil!—and then SQUEEZE to sever the clutch of organs in your grasp. DO
NOT PULL!!! It should be a decisive snip and if that’s not enough to separate the distal portions of the
sepals/petals, reposition forceps and take another bite. If you find yourself yanking on the

Figure10: Completing emasculation of flowers. The exposed pistil should be pollinated as
quickly as possible in order to reduce chances of inadvertent fertilization via air-borne
pollen grains.
flower you may as well abandon hope, she’s a goner. Repeat the process until you begin to gain access to
the stamens (remember, there are 6 of them). Ultimately your goal is to snip off the anthers from each,
easily done by clipping the thin shaft of the stamen. With patience and practice this can be done
routinely. [I apologize for all of the BOLD text and exclamation pointed warnings in the preceding, but it
took us a while to figure out how to avoid damaging flowers in the course of emasculation and I want to
spare you the experience. The bottom line is that there seem to be at least two key pitfalls: stretching
organs and stressing their confluence at the base, and avoiding any other damage to the pistil. Do either
of those to excess and you will almost certainly visit your flower in several days, filled with hope and
anticipation, only to find it withering away at the end of its pedicel.]
Now things get easy. Once you’ve (gently) removed outer organs and anthers you should be left
with an exposed pistil ready for pollination. Pollination for crosses is just like pollination for selfing except
the flower is taken from a different plant. Remove a flower from the intended father, squeeze to splay-
out anthers, and then dab pollen onto the pistil of the mother flower. Be sure to label the flower that
you’ve crossed onto. It’s tempting to think that you’ll be able to distinguish it from adjacent,
unemasculated flowers and that’s true—for about three days. After that, however, those other flowers
naturally lose their outer organs and come to look just like the one you crossed onto. We use colored
sewing thread to mark the pedicel of an out-crossed flower and different colors if multiple but distinct
crosses were performed on several flowers of a given plant. Take a 1 ½-2” piece of thread and tie a loose
overhand knot. Grasp the intertwined threads with your forceps, niggle it over the flower and down onto
the pedicel. Holding one end with your fingers and the other with the forceps cinch it up—obviously not
too tightly but tight enough that it won’t fray and fall off in a passing breeze.
Figure11: Use forceps to grasp the flower to be used for pollination at its base—the
organs will splay out to expose anthers (left). Remove the flower from its pedicel, invert,
and brush the pistil of an emasculated flower to leave a thorough coating of pollen
(center). Use colored thread tied loosely around the pedicel to demark the crossed flower
(right)
A couple of additional points are worth noting here. First, in some cases you may be able to
bypass emasculation altogether by pollinating exposed pistils that sometimes emerge from young
flowers. Whether it’s mere chance or some unknown variable in the growing environment, exposed
pistils sometimes do present themselves. If close inspection shows no signs of pollination, lathering on a
saturating coat of pollen seems a reasonable thing to do: the applied non-self pollen grains will have a
considerable head start over the self pollen grains released later and in any case should block access to
surface cells of the stigma. We’re not recommending this, we’re just sayin’…. Second, and somewhat
related, professional breeders typically ‘hood’ their crossed flowers to prevent unintended pollination by
either self pollen or any other source that may be in the vicinity. This is undoubtedly good practice but
probably overkill in most classroom situations for which we expect our line will have most use. Our
reasoning is as above—provided that you coat pistils with sufficient pollen from the intended donor, the
head start and occupancy achieved should be sufficient to minimize/eliminate the possibility of
unintended crosses.
Finally, it’s also worth considering the strategy of designing crosses to minimize or enable
detection of unintended matings. The biggest concern in most cases is harvesting seeds and growing
plants that result from inadvertent self-pollination. For example, if you were to take pollen from a
recessive mutant and use it in a cross to a wild type FPsc mother plant, the F1 plants would look wild
type regardless of whether the seeds were the result of the intended cross or an inadvertent self-
pollination. On the other hand, had we thought more strategically, it’s easy to tell the difference if the
cross was reversed: use pollen from wt FPsc to fertilize ovules of a homozygous mutant mother plant. If
the F1 are from the intended cross they’ll be wild type, if they’re from an inadvertent self of the mother
plant, they’ll be phenotypically mutant (obviously this only works if the mutant allele is recessive).
Sometimes this strategic decision is not practical—some mutant plants are feeble and largely incapable
of bearing seeds of any sufficient quality or quantity—so you have to keep issues regarding fecundity in
mind, as well.
7. Terminate the apical meristem and maintain plants until signs of maturation and senescence
become evident; diminish and then cut-off watering.
Terminate the apical meristem: This phrase means to remove any incipient flowers beyond
those that you reasonably hope will produce useful and non-crappy seeds. We have found that the FPsc
line can readily bear 8-12 fully fertilized seed pods (siliques), each holding 15-25 maturing seeds. If you’re
selfing to propagate a lineage (e.g., to advance an F1 plant to yield a segregating F2 seed family, or to
amplify stocks of true-breeding wild type or homozygous mutant lines) it’s best not to ask or allow your
plants to bear more than those numbers of progeny. Not only will your seed quality be diminished, but it
will take longer for the plants to reach senescence and for you to be able to harvest seeds and go on to
the next generation. Thus, if you’re propagating lineages through selfing, do hand pollinations at 3 and 5
or 6 days after the first flower opens, give the plant another day or two, and then use forceps to cleave
the remaining inflorescence buds from the previously fertilized flowers. If you are daring you can do so
immediately after the last round of hand-pollination, but we like to allow a bit of stem distance to
emerge that will separate the open stem wound resulting from termination to the last-fertilized flower. If
you’re doing crosses and are confident that the cross “took”—as evidenced by vigorous elongation of the
pistil/silique—let a few more flowers form, hand-pollinate those (always a good practice in genetics
experiments), and then terminate the meristem as above.
The rationale behind this recommended strategy is as follows: Plants (as with all organisms)
have a drive to survive. Upon transition from vegetative (leaf-forming) development to reproductive
(flower-forming) mode, individual plants will continue to flower as long as suitable conditions exist or
until the plant reaches a tipping point that balances the resource demands of growing embryos versus
new flower/embryo production. It is a well-known phenomenon in the plant research community that
male-sterile plants (e.g., those in selfing varieties unable to effect fertilization) flower FOREVER—or at
least it seems like forever in researchers’ minds—whereas fertile plants flower for some typical period of
time and then cease flower production. While the genetic and molecular details remain almost wholly
obscure, the inescapable conclusion is that there exists a feedback mechanism that signals to the plant
that some “normal” level of reproductive success has been achieved. Plants thereafter cease
reproductive development and, instead, allocate resources derived from photosynthesis to nurturing
growth and development of embryos and seeds. As experimental scientists, our estimation of the
“normal” level of reproductive success is driven not by innate tendencies of the plant, but by
experimental imperatives. By terminating the meristem, we send an unmistakable signal to the plant
that it’s time to stop flowering and to devote resources to raising healthy seeds. [If you are as obsessive
about hurrying through generations as we are, you might also take care to terminate secondary floral
inflorescences that either exist when you terminate the apical meristem, or emerge as a consequence of
releasing apical dominance through removal of the primary shoot.] We’re getting pretty deep into the
weeds of plant reproductive biology here, so let’s return to first principles: once you are confident that
the plant has produced and can support development of a good crop of mature seeds, try to prevent it
from devoting any more resources to flowering.
Maintain plants: Be sure to provide adequate water during this critical period but, as noted
above, don’t drown the plants. Inadequate watering is one of the surest ways to produce a crappy seed
lot. Finding that Goldilocks balance—not too wet and not too dry—isn’t as difficult to do as it may sound
as written here. It only takes some time, attention, and common sense. Depending on growing conditions
(planting density being most important since widely spaced plants grow taller), it may be necessary to
stake the plants to prevent lodging, or, to use the technical term, “falling over”. We use 10”-12” bamboo
barbeque skewers that we get from a local hardware, big box store or online sources. Poke the pointed
end into the soil near the base of the plant and secure the plant to the stake by using a paper-coated
wire twist-tie (the sort you get with loaves of bread). Bend the twist-tie into the shape of a V and position
the vertex behind the skewer. Grasping the twist-tie, stake, and stem of the plant just below the
inflorescence, use your other hand to put in a couple of twists. It doesn’t take much pressure to keep the
tie in place since small slivers of bamboo will give the tie a grip; be sure that you don’t strangle the plant.
Signs of maturation and senescence: As noted above, successful pollination/fertilization is
indicated by an elongating pistil. That lengthening should be evident by 3 days post-pollination and will
continue until the carpels and enclosed embryos/seeds (collectively, the silique) reach a length of 2”-3”
after ~2-3 weeks. During that time there will be a fattening of the silique that results from seed

development and enlargement. In time, the contours of seeds within the silique will become apparent. At
the end of those 2-3 weeks further elongation and swelling will stop and you might think the process is
finished, but it will take ~2 more weeks before seeds are fully mature and ready to harvest. During that
time (which can seem painfully long), energy reserves are synthesized and stored in cells of the embryo
and the mother plant directs formation of a tough and ultimately hard, red/brown seed coat. Nearing
the end of the mother plant life cycle, she mobilizes nutritional resources from leaf tissues, as evidenced
by wilting and loss of the formerly deep green pigmentation. It is likely that the only photosynthetically-
active tissues late in the plant life cycle are the carpels, so be sure to keep them close to the lights.
Ultimately, photosynthesis ceases in the carpels, as well, and they become successively paler and then
brown. That browning of siliques typically begins at the tip, or “beak” as it is known in botanical circles
and later spreads throughout the silique.
Figure12: Maturation and senescence of flowers and siliques (seed pods). Successful
pollination/fertilization is indicated by elongation and swelling of the pistil over time (left).
Developing seeds become evident 1-2 weeks after pollination (center) and
senescence/maturation is complete ~5 weeks after fertilization (right).
During these late stages of the plant life cycle, water needs are substantially reduced so it’s
advisable to cut back on watering-- don’t leave them parched, just a little thirsty. When plant tissues,
especially the carpels, are ~50% brown, you can stop watering altogether and seeds can be harvested
after another week or ten days. It may be helpful to keep in mind the natural life cycle of rapid-cycling B.
rapa varieties. Similar to the related and invasive garlic mustard plant that is the scourge of woodlands
throughout the upper Midwest, these plants are among the earliest to germinate and begin growing in
spring, well before the competition gets started. They thereby face less competition for resources
(especially light) and use that window of opportunity to rapidly progress through vegetative and then
reproductive development. Lengthening days and warmer temperatures are a signal that the hot
summer months are coming, as is a decrease in available moisture. The plants perceive these
environmental cues and respond by devoting all resources towards seed production and maturation. We
try to mimic those cues, at least with respect to water availability, in order to accelerate the seed
harvest.
There is another important reason to avoid excessive watering late in the FPsc life cycle. One of
the goals of the breeding program that lead to both the canonical WFP variety and our FPsc plants is “a
need for speed”. Time is a precious commodity in both classrooms and laboratories and we hoped that
by minimizing the generational (seed-to-seed) time, our teachers and students might be able to conduct
experiments in genetics that span multiple generations. One outcome of pursuing that goal is that
dormancy in our seed stocks is readily broken. That is, whereas seeds of some plant species can remain
dormant in soil through periods of wet and dry, hot and cold, for long periods of time—sometimes
decades—our seeds barely see the stop sign at the end of development. Excessive watering towards the
end of the FPsc life cycle can lead to the phenomenon of vivipary, wherein seeds actually germinate
within the siliques of senescent mother plants (see figure 4). Needless to say, that is a condition to be
avoided.
8. Harvest the seeds and either plant immediately or store seeds in cool, dry conditions. If you will be
storing the seeds or have more than you can use at the moment, label the seed packets in such a way
that you can track their genetic history later on.
Harvest the seeds: Once the siliques are fully brown and dry you can harvest the mature seeds
within. Try not to let the plants become too dry, as the siliques become prone to shattering and you may
lose the seeds that fall out as you manipulate the plants. If you’re harvesting seeds from just one or a
few siliques, a folded-up 4”x6” note card (see seed tray, figure 3) is sufficient: just crack the silique(s)
between your fingertips and let the seeds fall onto the tray. Once all seeds have been recovered, take a
look at them to see whether they are round and full or shriveled or otherwise crappy. If it’s the latter, you
may need to plant extra seeds to start the next generation, since the germination rate may be reduced.
Gently fold the note card lengthwise and pour the seeds into an appropriate receptacle. We use coin
envelopes (#2 or #3 Kraft stock), but anything that can keep them secure and dry during storage is fine.
Coin envelopes work well because they are stackable and provide plenty of space for labels. They also
‘breathe’, preventing accumulation of moisture or humidity that may result if seeds were harvested ‘a
little green’. Try to avoid harvesting and storing plant debris with your seeds; it can also be a source of
moisture that could promote precocious germination of seeds or growth of fungi and molds during
storage. Place seeds in a cool place for long term storage, preferably over desiccant in a sealed (e.g.,
Tupperware) container.
Labeling seed packets: Before describing our preferred system for recording details of seed
stocks held in storage, we need to be clear on one more bit of terminology—the notion of seed families.
It’s not a complicated idea, but we’ve found that it takes a while for many of our students and other
users to grasp its meaning. It’s simply a term used to reflect the fact that all of the seeds harvested from
a silique or whole plant should have the same mother and the same father and are thus siblings—they
constitute a family (sans mom and pop). Depending on the genetic constitution of the parents, the sibling
seeds in a family may be genetically uniform (if the parents were homozygous at the relevant locus/loci)
or they may differ if the parents were heterozygous at one or more loci. In either case, the seeds
constitute a single family, and just as we use surnames to designate most human families, each seed
family should get a unique identifier. It is tempting to use a descriptive and literal designation scheme—
e.g. “selfed seeds from albino segregant grown on the window sill in June, 2009”—but such a system
quickly spirals out of control if you want to be able to track the genetic provenance of seed stocks
through multiple generations of selfing or crosses.
We recommend a numerical system in which each seed family received or harvested is assigned
a serial number that is recorded on the seed packet or other container and entered into a spreadsheet
with a fuller description of genetic attributes. For example, let’s say that Scott and Rick send you seeds
harvested from an albino/+ heterozygote parent. ¼ are predicted to be +/+ , ½ to be alb/+, and ¼ to be
alb/alb homozygotes. Upon receipt of the seeds, you should assign to the seed family the next available
number in your stock list—let’s say it’s 103. You can add whatever additional information that you want
or will fit on your seed packet, but make sure that “103” is written at the top of the packet. So now, with
your heart racing and your mind spinning with anticipation of seeing what the plants will look like, you
plant them out. In short order, you will see that about ¼ of the resulting plants will be distinctly
yellow/pale green and ¾ will be just as green as the wild type FPsc plants (from your seed family stock
#1) that you grew as controls. Wow! you think, that’s a cool 3:1 ratio among plants in family #103;
maybe I can help my students to recognize that among the phenotypically wt sibs in that family, 2/3 are
heterozygous for the alb allele, so you have them self-pollinate all wt plants and collect the seeds. How
would you designate the plants and how will you designate the progeny seed families?
The individual plant designation is as simple as naming humans (although reversed): surname +
given name. Thus, sibling plants in the albino family 103 can be designated 103-1, 103-2, 103-3, etc.
When seeds produced by self-pollination of, say, plant 103-2 are harvested they, too, constitute a family
(although mom and dad were technically the same individual and that may be where students get
confused) and so merit a unique identifying number or surname (e.g., stock #104 derived from self-
pollination of plant 103-2). Ink and bytes are cheap, so you can add all of the descriptive verbiage that
you want in the hard-copy lab notebook or spreadsheet and some amount can be added to the seed
packet. The key to being able to trace the genetic history of your seed stocks, though, is a consistent
system of designating both individual plants and individual seed families.
9. Keep in mind that, since the FPsc variety has been extensively inbred, seeds produced by
true-breeding lines (whether wild type FPsc or homozygous mutants) will breed true via self-
pollination. The genetic uniformity of FPsc plants facilitates genetic analysis of mutant alleles since, in
segregating seed families, we intend that the only difference is the presence or absence of particular
allelic variants, whose effect on plant phenotype can be assessed in an otherwise uniform genetic
background. A practical benefit also ensues: provided that you and your students take care to keep
plants in optimal growing conditions through the life cycle, seeds harvested in excess of those needed
for immediate experiments can be used as starting materials for future implementation of classroom
exercises.
An important exception to this general rule applies to seed stocks distributed as FPsc/R500
hybrids. R500 is a naturally selfing, thoroughly inbred line of B. rapa that is grown in areas of India and
Nepal and whose seeds are used as a source of cooking oil or of condiments and spices. R500 was
selected as the polymorphic partner to FPsc in our resources. The polymorphic phenotypic attributes
range from the macroscopic (plant size, growth habit to molecular levels (R500 is the source of PCR
fragment length polymorphisms that we use in genetic mapping experiments. Most of the seed stocks
we distribute are in the form of F2 families derived from an initial FPsc mutant x R500 cross in which
mutant alleles from the FPsc parent will be segregating. However, it is important to keep in mind that
many (likely thousands) of allelic variants at other loci will also be segregating in such families and so
each F2 plant will have a unique genetic constitution. This genetic variation is essential in order to
extend activities and exercises from classical (Mendelian) to molecular genetic analysis but may
complicate or prevent propagation of seed lineages over time, since the derivative lines will not be true-
breeding.

10. Relax!!! The preceding is meant to provide a thorough guide to help you and your students
to make best use of the seeds and plants in your classroom, but we realize that the bolded terms and
scary warnings may be a bit daunting to the novice grower. There are really just a few common sense
principles to keep in mind:
-- Crappy seeds make crappy plants, crappy plants make crappy seeds. It pays to
select seeds that are full and nearly round when planting, and to provide ample soil and
spacing during plant growth and development.
-- Keep the soil moist but don’t drown the plants.
-- Do what you can to provide adequate light to support plant growth and
development.
--Bottom line: If you and your students take care of the plants, they’ll take care
of you.
11. Remember that help is just an email away. If you have questions, problems, or ideas for
new uses of these novel plant resources don’t hesitate to drop an email to Scott Woody
(swoody@wisc.edu) or Rick Amasino (Amasino@biochem.wisc.edu) at UW-Madison. We will do our best
to get back to you as soon as possible.
